I do not wish to pretend expertise in any of these fields, but rather to suspend my ignorance and try to find some connections between them in the meantime. I am writing this as an exploration, and I welcome and expect corrections on my explanations and speculations. However, I also welcome those who shy away from a paper as soon as mathematics makes its first appearance–my job is just to share my excitement about math and its connection with living things. I will begin by explaining fairly simple concepts, partially to keep the larger subject from slipping out from beneath me, but also to establish a method of building an understanding of the more complicated topics. Some of these topics are modeled after nature, elegant explanations of natural phenomena. Others are abstract: the math was put first and allowed to think freely.
Golden Spirals
To learn how to discover connections between mathematics and nature, I’ll begin with what is probably the most famous example: the golden spiral. Pretend for a moment that you’ve never seen or heard of golden spirals before, and that you don’t know anything about the golden ratio. A good question to ask yourself is “what makes this thing special?” The first observation you might make is that the golden spiral is self-similar. If you zoom in or out, nothing changes. You can imagine how you could get lost, not knowing how far in or out you are. But really, the golden spiral is just one of a larger set with that property, the logarithmic spirals.
Logarithmic spirals in nature exhibited in Aeonium tabuliforme leaves and a snail’s shell.Images: Wikimedia Commons, used under Creative Commons license
So, what makes this particular logarithmic spiral special? One answer could be that it isn’t–there are many non-golden logarithmic spirals in nature. In fact, the examples above certainly aren't exactly “golden”. But to faithfully attempt to answer the question we should try to precisely define a golden spiral, so here it is: a golden spiral is a logarithmic spiral with a growth factor of $ \frac{1+\sqrt{5}}{2} $.
Stay with me, and try not to worry about what exactly a growth factor of a logarithmic spiral is. For now we’d better think about what makes this specific number special. We call this number $\varphi$, “phi”, but where does it come from? Luckily there is a beautiful answer. Imagine two line segments, a and b. If you choose their lengths so that $ a:b = (a+b):a $, then $ \frac{a}{b}=\varphi $. In other words, try to find two numbers such that the ratio between them is the same as the ratio between their sum and the larger of the two.
If you turn those segments into squares $A$ and $B$, you can arrange them together to visualize how good your ratio is. Play around with different ratios and see if you can find gold. In this arrangement, the golden ratio manifests an open space below $B$ that is precisely the same shape as the rectangle surrounding both squares. Then, when the pattern repeats, it fills its own gap endlessly.
2.000
I must point out that this is only a stepwise approximation and not a true pure-bred logarithmic spiral, but I'll be controversial and say it doesn't matter for now.
I'm hoping you were able to approximate $\varphi$ yourself, but if not you can click on the hyperlink to a 3-digit approximation. When you scroll through these different ratios it becomes clear that the self-similarity of the golden spiral supersedes that of any other logarithmic spiral; it is the self-similarest! As promised, we now have a logarithmic spiral with a growth factor of $ \varphi $, but we also now have a real idea of what a logarithmic spiral’s growth factor is, which I think is more useful than a purely mathematical definition.
So, if golden spirals are particularly self-similar, we can rephrase our earlier question as “what is special about self-similarity?”. Imagine you have one of those inner rectangles with dimensions $ 1:\varphi $. You could then build outward with squares, creating that same pattern as before, forever. If you did have the infinite sequence of squares, any single square you chose would be identical. So, if we think of our placement of proportional squares as an algorithm for growth, it is now clear why this type of structure is so friendly to living things: a simple step can be followed over and over for controlled growth in all directions.
Cells as Machines
That line of reasoning is very interesting, but equally dangerous. Obviously there is no real "growth algorithm" that living things follow, so before we continue down this road we should recognize its failures. Mechanistic characterization of biology is extremely common, and it lives at the center of biology’s most active and most funded fields such as cancer research (Hanahan et al. 2000). This idea has issues beyond oversimplification, it mischaracterizes biological components from the ground up, leading to a framework of studying biology that is unproductive and misleading (Nicholson 2019).
While mathematical biology–the attempt to model living things with mathematical formulas and models–might seem to be an even more extreme case of this fallacy, it is important to pinpoint the specific issue with the machine conception of the cell. I’ll give you a hint: it certainly isn’t simplification. Machines are defined by their parts and how they come together; they can be constructed faithfully with a set of instructions. Machines are highly specialized and constrained in their performance. Machines may also be interrupted, taken apart, put back together, and they should continue functioning as before.
The contradiction between cells and machines as a metaphor can be boiled down to two traits of cells, their self-organization and continuous undefined function (Nicholson 2019). Additionally, I would argue that a cell cannot be constructed faithfully with a set of instructions. Certainly you’d need more instructions than just the sequence of DNA.
Mathematical biology certainly has a responsibility to be cautious of these types of fallacies, but it seems to be a promising way out of this misdirection. Our discovery about golden spirals wasn’t really about how living things work at all, it was the characterization of a form (a good thing) and then conjuring up an anthropocentric explanation of it (not so good). Idealization is a clear necessity to science, but it must be done cautiously and for the sake of discovery. When those ideals are too central to our actual perceptions our research can meander down suboptimal paths for decades on end.
Bifurcation
The logistic map is another example of mathematics that is central to biology, and can be looked at in quite the same way as we analyzed golden spirals. Take a population with some rate of reproduction proportional to the population size, and some rate of starvation proportional to the population’s carrying capacity. To find a population $ x_{n+1} $ as a proportion of carrying capacity, you could multiply the previous iteration’s population by some growth rate, $ r $, and constrain the maximum possible population with the term $ (1-x) $. This equation, $ x_{n+1} = rx_n(1-x_n) $, is the logistic map. And with different growth rates and initial populations, you can watch as populations evolve over time and settle on equilibria (May 1976). Initial population $x_0$ has no effect on the end behaviors of a population, so for our purposes we can select $ x_0 = 0.25 $ for consistency. So, let's investigate the equilibria for a few values of $r$.
r = 3.00
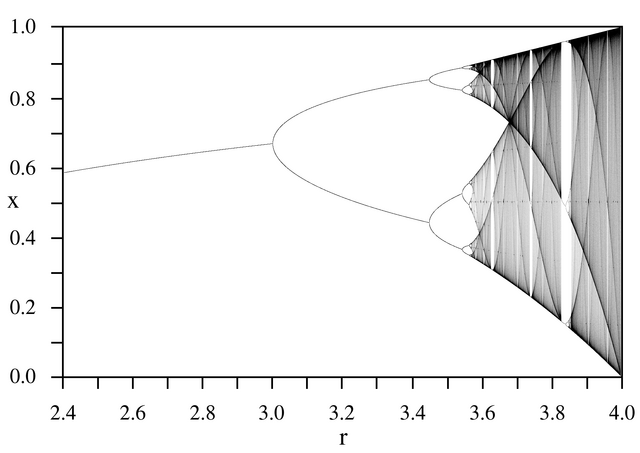
At $r=1$ the growth rate is so small that the population just fizzles out. At $r=2$ it quickly settles at $x=0.5$, and stays there iteration after iteration. At $r=3.1$ there is an interesting balance between growth and death, and iterations bounce back and forth between two values. Then, at $r=3.47$ there is another bifurcation, and there are 4 relatively stable values that the population bounces around between predictably. But even just $0.1$ units higher, at $r=3.57$, that stability begins to dissolve. And at $r=4$ the pattern is completely lost, and there is no equilibrium at all. In fact, early computers used to use this function to generate pseudorandom numbers. Critically, though, this data will always be the same no matter how many times you run the simulation–these numbers are not actually random at all. I strongly recommend that you play around with this, scrub through the values and try to find some interesting behaviors. There is clearly something interesting going on with equilibria in the logistic map equation, so let’s try plotting the numbers that $x$ “settles” on against different values of $r$.
The logistic map, $x_0=0.25, r\in(2.4, 4.0)$.Image: Wikimedia Commons, used under Creative Commons license
This is the logistic map, and it is a fractal pattern riddled with areas of seemingly random complexity and sudden order. Comparing this to our data, we see a single equilibrium below $r=3$, and then two equilibria until $r=3.5$, where the chaos begins. Note that later, around $r=3.83$, we once again find some stability. What is going on with that?
A good question to ask is if we can predict when this map splits into two. In 1975 the mathematical physicist Mitchell Feigenbaum discovered that these bifurcations occur more and more rapidly, each one coming approximately 4.669 times sooner than the last.
The logistic map, and chaotic bifurcation patterns in general, exist as clearly in nature as the golden spiral. In real organism populations as well as thermodynamics, fluid dynamics, and cardiology there have been clearly demonstrated bifurcation patterns that lead to chaotic behavior, much like the logistic map (Garfinkel et al. 1992, Libchaber et al. 1982, Shaw 1984). For example, in a famous experiment it was shown that as a rabbit heart undergoes fibrillation, there is a period doubling in heartbeat that eventually leads to death. Using chaos theory, researchers determined optimal times to apply electrical shocks to the rabbits, returning their heartbeats to periodicity with some success (Garfinkel et al. 1992).
Patterns
Conway’s Game of Life
I want to show you a different type of simulation, something that was described by its creator as a "zero player game." It can be described with three rules (different rule sets, often with four rules, describe the same game in slightly different ways, so if these are not intuitive the other explanations may be helpful):
A living cell with less than two neighbors dies (starvation)
A living cell with more than three neighbors dies (overpopulation)
A dead cell with three neighbors comes to life (reproduction)
This game is extremely computationally cheap, unlike reaction-diffusion, and you can play it yourself quite easily with graph paper. This simple game yields surprising complexity, such as gliders that move across the array forever, or growing complexes that never stop.
See if you can create something that will sustain itself. Some formations will oscillate between two states eternally, while others will span across the cosmos creating more and more explorers, bound for the abyss. Some, eerily, remain completely still–not dead as far as the rules of the game are concerned, but lifeless, dormant.
Games like these are called "cellular automata," and there are endless examples of their fascinating behaviors, but rather than dive down that rabbit hole I want to make a connection to something a lot less intuitive than our simple rule set. Remember, living things don't like to follow rules.
Reaction-Diffusion
Alan Turing was a mathematician and computer scientist famous for his contributions to cryptography and historic role in World War II. But two years before his death, he wrote an important paper in the field of mathematical biology, “The Chemical Basis of Morphogenesis”. He was inspired by D’Arcy Thompson’s book, On Growth and Form, which connected mathematics to evolution, and argued for consideration of physical laws in the field of biology as a whole. Turing wondered if the same thing could be done with chemistry, if there were principles of chemical reactions that govern how organisms form (Ball 2015).
He describes a simplified embryo containing two morphogens, substances with varying physical and chemical properties that interact to determine the physical form of an organism. This definition is intentionally vague, and encapsulates genes, hormones, and even skin pigments (Turing 1952). He does well to describe the long list of limitations of this simplification, but carries on describing how certain properties of these morphogens could result in novel reaction diffusion patterns. The mathematics behind this type of interaction is precise, but quite the opposite to the logistic map in that the visual is more intuitive than the equations.
Reaction-diffusion simulation using the Gray-Scott model with $r=0.6$ and $k=0.6$.Visualized using RD Tool (Sims 2022).
Soak in these images, you can intuitively see how the black substance grows and spreads, folds inwards and oozes outwards. This particular simulation uses the Gray-Scott model, an iteration of Turing’s equations developed in the Chemical Basis of Morphogenesis. Turing's model is more general and versatile, capable of producing a wider range of patterns. The Gray-Scott model, being a simplification, is computationally less expensive and has been widely used for simulating pattern formation in reaction-diffusion systems.
Turing developed the following partial differential equations:
$$ \frac{\partial u}{\partial t} = D_u \nabla^2 u + f(u,v) $$
$$ \frac{\partial v}{\partial t} = D_v \nabla^2 v + g(u,v) $$
Despite their complicated appearance, they're actually quite simple. Take a distribution of a substance, smudge it around a bit, and then either take some away or add some more. There are two equations because there are two substances, $u$ is an autocatalyst (self-producer) and $v$ is an inhibitor. The functions $f$ and $g$ determine how the neighboring concentration of one substance influences the other, and the laplacian ($\nabla^2$) terms are similar to a convolution (like a smudging) of a particle of the substance, with the result dependent on the neighboring particles’ identities. The Gray-Scott model distinguishes itself by the chosen behavior of the autocatalyst and inhibitor. In Turing’s equations, the autocatalyst promotes growth of both substances, and the inhibitor inhibits the growth of both substances. In the Gray-Scott model the autocatalyst promotes its own growth, consuming the inhibitor. The inhibitor is added at rate r and the autocatalyst must be removed at rate k to keep the reaction going. The mathematics look quite similar:
$$ \frac{\partial u}{\partial t} = D_u \nabla^2 u - uv^2 + f(1-u) $$
$$ \frac{\partial v}{\partial t} = D_v \nabla^2 v + uv^2 - (f+k)v $$
The values of r and k can easily be altered to drastically change the behavior of the system, making these equations more approachable to build an understanding.
I strongly urge you to play around with Karl Sims' simulation of the Gray-Scott model, RD Tool. I would have embedded a simulation here, but to be completely honest I couldn't have done nearly as good of a job, his simulation is a true work of art.
Turing Completeness
A Turing machine is an abstract computer that consists of an infinite tape, a tape head that reads and writes symbols on the tape, and a set of rules that dictate its operations. The machine can move the tape left or right, change the symbol at the current position, and change its state based on the rules. This machine, despite its simplicity, can compute any algorithm, no matter how complex. If any system can simulate this Turing machine, it is called “Turing complete”, and clearly if you can simulate a machine that can compute any algorithm, you can compute any algorithm.
Reaction-diffusion and the Game of Life differ in that reaction-diffusion reaches a sort of equilibrium, even if the substances never stop moving or interacting. Conway’s Game of Life is different, and you may find that on the millionth iteration the entire infinite plane of cells fizzles out and dies completely. The Game of Life is known to be a “Turing complete” system. This property of the Game of Life was demonstrated by Paul Rendell when he created a pattern known as a "universal constructor" that can be used to simulate any arbitrary cellular automaton, making the Game of Life Turing complete (Rendell 2011). To grossly simplify what this means, the Game of Life can be used to simulate any other Turing machine, which is a simple yet powerful abstract computer. Reaction-diffusion on the other hand is not known to be Turing complete. This hasn’t been proved one way or another, but the stability and predictability of reaction-diffusion is characteristically unlike a Turing machine.
Turing completeness is particularly important because of how it relates to our machine conception of the cell. Recall that the two traits of cells that make them non-mechanistic are their self-organization and continuous, undefined function. A Turing complete machine has a very defined function, and runs on iterations of instructions, not continuously. What makes reaction-diffusion promising isn’t that it might be Turing complete, it’s that it might actually be Turing incomplete. Reaction-diffusion is a continuous function, despite being computed in steps. If it was possible to compute with reaction-diffusion, it would be more biological than the computation of Turing machines (although I must admit, who knows what “more biological” even means in this context?).
Conclusion
The pattern of thought we first used to understand golden spirals–beginning with understanding the behavior of the mathematics at the simplest possible level, and then exploring the edge cases, where the definitions fail or bleed into others, or where properties emerge and fade away–has useful application in the exploration of other mathematical patterns that are exhibited by living things. While some of these patterns clearly exhibit order, many are chaotic with complex behavior. And yet, the field of chaos theory and other seemingly obscure topics in mathematics bring clarity to these situations.
Impressive progress in the field of mathematical biology has come from preliminary exploration of other fields, such as physics with Thompson or chemistry with Turing. Surprisingly, there don’t seem to have been any real breakthroughs in the past few decades, which to me is a sign that the field is ripe for another expansion into topics in chaos theory, computer science, modern physics, modern mathematics, and so on. I see this as the most promising frontier in modern biology, and yet it is largely out of the limelight of funding and research efforts.
Acknowledgements
I would like to sincerely thank Dr. Michael Gustin, Dr. Kathleen Beckingham, and Miles Gantcher for their insight and help proofreading this paper.
References
Rendell P. A Universal Turing Machine in Conway’s Game of Life. In: 2011 International Conference on High Performance Computing & Simulation. IEEE; 2011:764-772. doi:10.1109/HPCSim.2011.5999906
May RM. Chaos and the dynamics of biological populations. Nuclear Physics B - Proceedings Supplements. 1987;2:225-245. doi:10.1016/0920-5632(87)90020-X
Ball P. Forging patterns and making waves from biology to geology: a commentary on Turing (1952) ‘The chemical basis of morphogenesis.’ Phil Trans R Soc B. 2015;370(1666):20140218. doi:10.1098/rstb.2014.0218
Nicholson DJ. Is the cell really a machine? Journal of Theoretical Biology. 2019;477:108-126. doi:10.1016/j.jtbi.2019.06.002
Libchaber A, Laroche C, Fauve S. Period doubling cascade in mercury, a quantitative measurement. J Phyique Lett. 1982;43(7):211-216. doi:10.1051/jphyslet:01982004307021100
May RM. Simple mathematical models with very complicated dynamics. Nature. 1976;261(5560):459-467. doi:10.1038/261459a0
Sims K. RD Tool. https://www.karlsims.com/. 2022.
Turing A. The chemical basis of morphogenesis. Phil Trans R Soc Lond B. 1952;237(641):37-72. doi:10.1098/rstb.1952.0012
Shaw R. The Dripping Faucet as a Model Chaotic System. Aerial Press; 1984.
Hanahan D, Weinberg RA. The Hallmarks of Cancer. Cell. 2000;100(1):57-70. doi:10.1016/S0092-8674(00)81683-9